
Revista del Museo de Antropología
versión impresa ISSN 1852-060X versión On-line ISSN 1852-4826
Rev. Mus. Antropol. vol.14 no.3 Córdoba dic. 2021
http://doi.org/10.31048/1852.4826.v15.n1.34530
ARQUEOLOGÍA
Pastoreo en la Puna post-hispánica, zooarqueología del sitio Antigal laguna (Barrancas, Jujuy)
Pastoralism in the post-hispanic Puna, zooarchaeology of the Antigal Laguna site (Barrancas, Jujuy)
Julia Merler Carbajo*
*CONICET, Instituto de Biodiversidad y Biología Experimental, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Argentina. E-mail: julimerler@yahoo.com.ar
Resumen
Junto con los conquistadores europeos arribaron al continente americano diversas especies de animales domésticos, las cuales se dispersaron rápidamente por el territorio. En el Noroeste Argentino su incorporación a la economía local fue relativamente rápida y de gran importancia en el marco del sistema socioeconómico impuesto por los conquistadores. Esto fue así particularmente en la Puna, donde la preexistencia de prácticas pastoriles y de manejo de animales de rebaño representó una ventaja a la hora de incorporar estas especies y favoreció la participación indígena en el nuevo sistema económico. Sin embargo, la información disponible acerca del ingreso de estos animales al pastoreo tradicional andino, su manejo y los usos que se le dieron en esta región es escasa. Este trabajo presenta los resultados del análisis de los restos zooarqueológicos provenientes del sitio Antigal Laguna, ubicado en la Localidad Arqueológica de Barrancas (departamento de Cochinoca, provincia de Jujuy), sitio que da cuenta de la ocupación del área durante momentos post-hispánicos y presenta restos de fauna europea, lo que lo convierte en una fuente valiosa de información para conocer cuál fue el vínculo entre las poblaciones locales y estos animales entre el siglo XVII y XVIII.
Palabras clave: Noroeste argentino; Puna jujeña; Fauna europea; Arqueología histórica; Pastoreo.
Abstract
Along with the European conquerors, various species of domestic animals arrived on the American continent and quickly dispersed throughout the territory. In the Argentine Northwest, their incorporation into the local economy was relatively rapid and of great importance within the socioeconomic system imposed by Europeans. This was particularly true in the case of the Puna region, where the preexistence of pastoral practices and the management of herd animals represented an advantage when incorporating these species and favored indigenous participation in the new economic system. However, the information available about the incorporation of these animals into traditional Andean grazing, their management and the uses that were given to them in this region is scarce. This paper presents the results of the analysis of the zooarchaeological remains from the Antigal Laguna site, located in the Archaeological Locality of Barrancas (department of Cochinoca, province of Jujuy). This site accounts for the occupation of the area during post-Hispanic times and presents remains of European fauna, which makes it a valuable source of information to learn about the link between local populations and these animals between the seventeenth and eighteenth centuries.
Keywords: Argentinean northwest; Puna of Jujuy; European fauna; Historic archaeology; Pastoralism.
Recibido 18-08-2021
Recibido con correcciones 17-09-2021
Aceptado 09-10-2021
Introducción
La llegada de los conquistadores al Noroeste Argentino (NOA) en el año 1534 d.C implicó la imposición de un nuevo sistema económico y social, el cual se estructuraba principalmente sobre tres ejes: la minería, la ganadería y el comercio (Conti y Sica, 2011; Gil Montero, 2009; Palomeque, 2000). Sin embargo, la imposición y asimilación de este sistema no se dio de igual manera en todo el NOA, dependiendo fuertemente de la variabilidad ambiental y cultural de cada región (Noli, 1998; Sica, 2005). En el caso de la Puna, el hecho de que se trate de un área de difícil acceso y con un clima sumamente árido y poco apto para el desarrollo de la agricultura, determinó que la ocupación efectiva por parte de los españoles se diera tardíamente, a la vez que el control político fue menos estricto que el que se dio en otras zonas del NOA como la quebrada de Humahuaca (Aramendi, 2016; Palomeque, 2000).
En muchos casos, los grupos indígenas que habitaban la Puna pudieron mantener cierta continuidad en las practicas económico-culturales luego de la conquista (Palomeque, 2000). De hecho, un factor clave en la adecuación al nuevo sistema económico por parte de estos grupos, fue la preexistencia del pastoreo como actividad económica principal desde momentos prehispánicos, y la presencia de una amplia red de intercambio a través de caravanas de llamas. Estas dos prácticas constituyeron una ventaja a la hora de ingresar en los nuevos sistemas de intercambio mediante el sistema de arriería y facilitó la incorporación de la fauna doméstica traída por los conquistadores (principalmente cabras, ovejas, vacas y burros). Estos factores facilitaron la participación de los grupos indígenas en el sistema comercial y económico colonial. Las especies europeas, de acuerdo con lo planteado por las fuentes históricas, se dispersaron e incorporaron rápidamente a la cotidianeidad de los pueblos del NOA (Conti y Sica, 2011; Gil Montero, 2009) y continúan siendo parte de los rebaños de los pastores de la Puna en la actualidad.
Sin embargo, hasta el momento no se cuenta con demasiada información acerca del impacto que tuvo la conquista en la región de la Puna, particularmente en la estructura del pastoreo tradicional andino y las implicancias de la introducción de especies europeas en los rebaños. Esto se debe, por un lado, a la poca disponibilidad de documentos y fuentes históricas que hagan referencia a áreas de difícil acceso y aisladas como la Puna (Vaquer, 2016). Por otro, no hay demasiada presencia de fauna exótica en el registro arqueológico. Hasta el momento se han hallado restos en contextos domésticos (Nielsen et al. 2015; Quiroga, 2015), rituales (Capparelli et al. 2005), en entierros (Otero y Rivolta, 2015), en contextos vinculados con la minería (Angiorama et al. 2015; Vaquer, 2016), o utilizados como materia prima (Von Rosen, 1957). A esto se suma que no hay demasiados trabajos arqueológicos que hayan abordado la conformación de la estructura del pastoreo durante momentos posteriores a la conquista y las consecuencias de la incorporación de especies exóticas.
En este trabajo se presentan los resultados del análisis del material faunístico del sitio arqueológico Antigal Laguna, ubicado en la Localidad Arqueológica de Barrancas (provincia de Jujuy). Este sitio presenta fauna europea y material arqueológico de cronología histórica, así como con dos fechados que enmarcan la ocupación entre los siglos XVII y XVIII. Además, sus características arquitectónicas y ubicación permiten pensar que se trata de una base residencial o “Casa de Campo”, las cuales forman parte del patrón de asentamiento de los pastores andinos y suelen ocuparse durante la estación húmeda (Madero, 2000; Yacobaccio, 2007). Es por ello que este sitio constituye una fuente de información sumamente valiosa a la hora de abordar el proceso de incorporación de especies domésticas europeas en la Puna y en el sistema pastoril tradicional andino.
Área de estudio
La Localidad Arqueológica de Barrancas se encuentra ubicada en el departamento de Cochinoca, provincia de Jujuy, al noroeste de las Salinas Grandes y a 3600 msnm, dentro de la ecorregión conocida como Puna de Atacama. Esta presenta las características climáticas y ambientales de un desierto de altura, con alta radiación solar, baja presión atmosférica y precipitaciones estacionales, lo que determina una distribución heterogénea de la vegetación en pisos altitudinales (Cabrera, 1976; Morales et al., 2018).
La localidad se extiende a lo largo de 8 km en dirección norte-sur sobre la planicie aluvial y terrazas fluviales del río Barrancas, el cual está flanqueado por altos farallones de ignimbrita. A lo largo del valle de este río se identificaron 41 sitios arqueológicos que se distribuyen sobre ambas márgenes del río y se encuentran emplazados tanto sobre las terrazas como en las quebradas tributarias. La enorme diversidad que presentan los sitios (sitios habitacionales, aleros, sitios con estructuras, etc.) y el hecho de que muchos de ellos se encuentren asociados con gran cantidad de paneles con representaciones rupestres conllevó a la declaración del área como Reserva Natural y Cultural Municipal.
De acuerdo a las investigaciones arqueológicas realizadas hasta el momento, hacia los 3430 años AP se observa un momento de transición entre ocupaciones cazadoras-recolectoras y las últimas etapas de la domesticación de camélidos. Las primeras evidencias claras de pastoreo de estos animales se registran hacia los 3200 años AP, junto con el intercambio mediante caravaneo. Esto último se observa principalmente a partir de la presencia de motivos rupestres y por la presencia de bienes exóticos en el registro arqueológico (Yacobaccio et al., 2018).
A lo largo de la cuenca del río Barrancas se pudieron registrar cuatro sitios arqueológicos que dan cuenta de ocupaciones durante momentos posthispánicos en la quebrada: Antigal Laguna, Alero del Caballo, Alero de las Cruces y Alero de los Caballos. Estos sitios cuentan con material arqueológico de cronología hispana, como cuentas de vidrio y mayólica, y con fechados que ubican las ocupaciones alrededor de los siglos XVI y XVIII. Los tres últimos se encuentran, asociados con representaciones rupestres de motivos hispánicos, tales como jinetes, caballos y cruces cristianas. Antigal Laguna y Alero de las Cruces son los únicos sitios que hasta el momento presentan restos de fauna europea.
En la actualidad, en el valle se observa una continuidad de las prácticas de pastoreo, tanto de camélidos (Lama glama) como de especies introducidas (Capra hircus, Ovis aries, Bos taurus y Equus asinus), las cuales conforman una parte estructural de los rebaños. Esto se observa no solo en esta localidad, sino en toda la Puna seca, siendo la estructura mixta de los rebaños un elemento clave en la organización de la dinámica del pastoreo y a la hora de hacer frente a situaciones de riesgo ambiental (Madero, 2000; Yacobaccio, 2007).
Sitio arqueológico Antigal Laguna
Antigal Laguna se encuentra ubicado sobre la margen izquierda del río Barrancas, en el sector medio de la quebrada, a 23º 18’ 06’’ S y 66º 05’ 18,5’ O (Figura 1). El sitio está compuesto por tres estructuras, dos corrales y cuatro trojas, distribuidos a lo largo de, por lo menos, 150 m lineales en una terraza estructural adyacente a paredones bajos de ignimbrita. Este sitio fue abordado en el año 2014, en el cual se llevaron a cabo prospecciones superficiales y subsuperficiales. La primera implicó una recolección de material superficial en el área, dentro y fuera de las tres estructuras presentes en el sector. Se relevó un área total de 3205 m2 en la cual se recolectaron diversos artefactos de material lítico, cerámico, óseo y metálico.
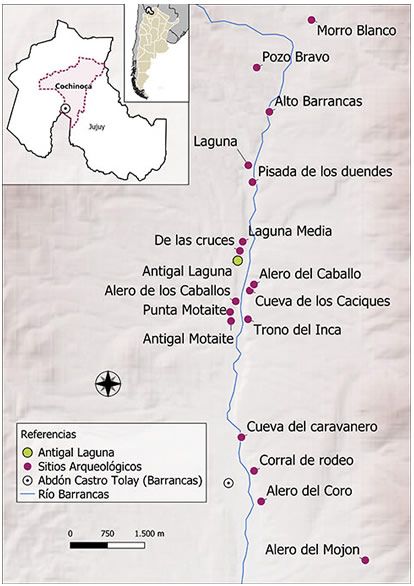
Figura 1. Ubicación de la Localidad Arqueológica de Barrancas y del sitio arqueológico Antigal Laguna.
Figure 1. Localization of Barrancas Archaeological Locality and Antigal Laguna archaeological site.
La prospección subsuperficial, por otro lado, constó de dos sondeos de 1 m2, uno en el interior y uno en el exterior de la estructura 3, adyacentes a la pared suroeste de la misma. La excavación se realizó a partir de las capas naturales, siguiendo la coloración y textura del sedimento, lo que permitió determinar la existencia de dos capas, tanto en el interior como en el exterior de la estructura. Es relevante mencionar que en la cuadrícula externa ambas capas se encuentran separadas por un derrumbe de rocas, posiblemente de la pared de la estructura. Esta cuadrícula tiene una profundidad de 61cm y brindó una gran cantidad de material faunístico, así como artefactos líticos y tiestos cerámicos. En la capa dos, se identificaron cuatro lentes de ceniza superpuestos, uno de los cuales, el que se encuentra por encima, brindó el fechado sobre carbón de 200±20 años AP (GifA #17281 / ECHo #1711 200±20BP) (Figura 2a). La estructura 3 cuenta además con otro fechado de cronología similar, también sobre carbón (GifA 20063: 200±23 AP, AMS).
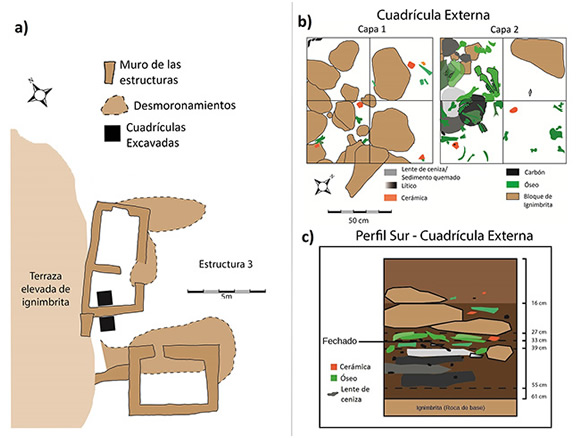
Figura 2. a) Estructura 3 y ubicación de las cuadrículas excavadas. b) Planta de excavación capas 1 y 2 de la cuadrícula externa. c) Perfil sur de la cuadrícula externa.
Figure 2. a) Structure 3 and location of the excavated grids. b) Excavation plant layers 1 and 2 of the external grid. c) South profile of the external grid.
La cuadrícula interna tiene una profundidad de aproximadamente 60 cm y presenta tres estructuras de combustión correspondientes a lentes de ceniza en la capa 2. Se obtuvo material cerámico, cuentas y fragmentos de vidrio, junto con un ojo de aguja confeccionada sobre materia prima ósea (Figura 3b). El material faunístico obtenido de esta cuadrícula y de la cuadrícula externa conforma la muestra analizada en este trabajo.

Figura 3. Material arqueológico proveniente de la excavación. a) Cerámica proveniente de la cuadrícula externa. b) Fragmentos de vidrio, ojo de aguja ósea, cuentas de vidrio y cerámica provenientes de la cuadrícula interna. Escala 10 cm.
Figure 3. Archaeological material from the excavation. a) Ceramic from the external grid. b) Glass fragments, bone needle eye, glass beads and ceramics from the internal grid. Scale 10 cm.
Muestra y metodología
El abordaje de la problemática planteada en este trabajo se realizó a partir del análisis de los restos faunísticos provenientes del sitio arqueológico Antigal Laguna. Con respecto a los antecedentes históricos, no se consultaron fuentes primarias. La información fue obtenida de fuentes secundarias y trabajos realizados por historiadores.
La muestra se dividió en cuatro unidades de análisis correspondientes a las capas 1 y 2 de la cuadrícula externa y la interna. Esto se realizó de manera de ponderar las relaciones espacio-temporales entre los conjuntos y evaluar los procesos de formación de sitio y del registro.
Identificación taxonómica y anatómica
Se llevó a cabo la identificación taxonómica y anatómica de los especímenes, a partir de la utilización de manuales osteológicos (Adams y Crabtree, 2012; Hopkins et al., 1970; Pacheco Torres et al., 1986) y colecciones comparativas pertenecientes al Instituto de Arqueología (Facultad de Filosofía y Letras, UBA) y al Museo de Ciencias Naturales Bernardino Rivadavia. Sobre esta base se obtuvieron distintas medidas de cuantificación tales como el Número de Especímenes Identificados por Taxón (NISP), Mínimo Número de Elementos (MNE), Mínimo Número de Unidades Anatómicas (MAU y MAU calculado en porcentaje) y el Mínimo Número de Individuos (MNI) (Lyman, 1994; Mengoni Goñalons, 1999).
A su vez, se calcularon los grupos etarios del taxón Camelidae a partir de la erupción y desgaste dentario (Wheeler, 1982) y de la fusión de las epífisis (Mengoni Goñalons, 1999). Para el resto de las especies esto no fue posible, dada la ausencia de las partes diagnósticas necesarias para determinar la edad. La única excepción corresponde a un metapodio de Bos taurus para el cual pudo identificarse la fusión de la epífisis (Hongo, 1998).
Procesos tafonómicos
A continuación, se abordaron variables que permitieran evaluar la incidencia de agentes tafonómicos sobre los especímenes. Se midió la meteorización mediante la escala ordinal planteada por Behrensmeyer (1978) y se consideró la presencia de marcas de carnívoro, roedores y raíces sobre la superficie ósea (Binford, 1981; Lyman, 1994; Mengoni Goñalons, 1999), así como la depositación de manganeso (Barrientos et al., 2007) y carbonato de calcio (Cornaglia Fernandez, 2012). Las marcas de carnívoro se clasificaron de acuerdo a los tipos planteados por Binford (1981): pozos, hoyos, mascado, borde aserrado, ranurado y ahuecado.
Evidencias de acción antrópica
Para evaluar la asociación de los conjuntos con actividades antrópicas, se consideró la presencia, tipo, distribución en la topografía del hueso y orientación de marcas antrópicas (Binford, 1981). Se evaluó también la presencia de fractura transversal, espiral y longitudinal y su asociación con marcas de percusión y/o de carnívoro (Mengoni Goñalons, 1980). Por último, se consideró la presencia de termoalteración, clasificando los especímenes en No quemado, Quemado, Carbonizado y Calcinado (Mengoni Goñalons, 1999).
Marco de referencia etnoarqueológico
Las evidencias disponibles hasta el momento sugieren que el pastoreo se instaura en la Puna como actividad económica de relevancia aproximadamente entre los 3000 y 3500 años AP (Olivera, 1997; Samec, 2014, entre otros). Esta experiencia previa en lo que hace al manejo de animales domésticos de rebaño permitió una rápida incorporación de las especies introducidas por los españoles (principalmente cabras y ovejas) y una diversificación en la actividad ganadera (Conti y Sica, 2011; Gil Montero, 2009).
La estructura del pastoreo andino actual está fuertemente condicionada por la posibilidad de aprovechamiento de pasturas en un ambiente donde los recursos se distribuyen en parches. Esto determina la organización mediante un sistema de movilidad de los rebaños y pastores entre distintas franjas altitudinales durante las estaciones húmeda y seca (Browman, 1987; Yacobaccio, 2007; Yacobaccio, 2014). Hoy en día, las especies domésticas europeas constituyen una parte estructural del sistema pastoril andino y de la cotidianeidad de los grupos humanos que habitan la Puna jujeña. Diversos trabajos etnoarqueológicos dan cuenta de los usos que se le da a estas especies, qué requerimientos plantean a la estructura del pastoreo y cómo estos se articulan con el manejo de las especies nativas (Lama glama) (Madero, 2000; Yacobaccio, 2007, entre otros).
Estos estudios constituyen un marco de referencia sumamente valioso a la hora de plantear hipótesis sobre el tipo de manejo utilizado tanto para los camélidos como para las especies introducidas durante momentos post hispánicos, sobre todo al articularlos con la evidencia arqueológica disponible y la información proveniente de fuentes históricas. Los animales que más se adaptaron al sistema pastoril andino fueron las ovejas y las cabras, las cuales predominaban en los rebaños frente al ganado vacuno (Aramendi, 2016; Noli, 1998). Actualmente su uso está principalmente destinado al consumo doméstico, al igual que los camélidos (Yacobaccio, 2007), algo que también se menciona para momentos históricos en las fuentes (Assadourian, 1982) y en el registro arqueológico local (Nielsen et al., 2015). A su vez, la diversidad de los rebaños se constituye como clave a la hora de hacer frente a situaciones de riesgo ambiental, modificando los patrones de matanza de manera de poder, en condiciones climáticas adversas, seguir explotando las distintas especies y, al mismo tiempo, garantizar la viabilidad productiva de los rebaños (Madero, 2000).
Durante momentos históricos las vacas se criaban en mayor medida en las zonas bajas y su paso por la puna estaba dado principalmente a través de las redes de arriería y abastecimiento de los centros mineros (Aramendi, 2016). Hoy en día tampoco se crían en grandes cantidades y su presencia está mas vinculada a cuestiones de status más que a consumo (Göebel, 2002; Rabey et al., 2008).
Resultados
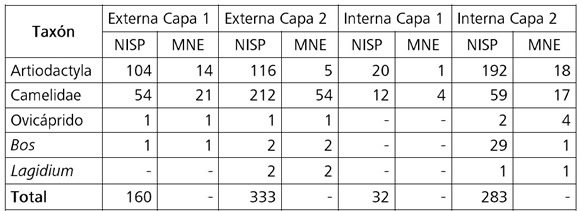
La muestra está compuesta por un total de 1714 especímenes óseos, de los cuales 808 (47,1%) pudieron ser identificados a nivel taxonómico. Los 906 especímenes restantes fueron categorizados como Indeterminados (Indet). Los taxones identificados incluyen la familia Camelidae (41,7%) y los géneros Bos (4%) y Lagidium (0,4%). Los especímenes pertenecientes a los géneros Capra y Ovis (0,5%) fueron denominados bajo el término “Ovicáprido”, dada la ausencia de partes diagnósticas que permitieran diferenciar entre ambos. Por último, aquellos especímenes que no pudieron ser adscriptos a una categoría taxonómica más específica se agruparon bajo la categoría de Artiodactyla. Estos son principalmente fragmentos de tejido esponjoso y de huesos largos carentes de rasgos diagnósticos, y corresponden al 53,5% de los especímenes identificados. Las proporciones en que están representados los distintos taxones se repiten en todos los conjuntos analizados (Tabla 1).
Tabla 1. Composición taxonómica (NISP) y anatómica (MNE) de la muestra.
Table 1. Taxonomic (NISP) and anatomical (MNE) composition of the sample.
Con respecto a la representación de partes esqueletarias, de acuerdo con el MNE de Camelidae se observa una representación muy pareja del esqueleto axial y el apendicular (Tabla 2). Si bien de acuerdo con el MAU% los elementos del esqueleto axial suelen están representados en porcentajes bastante más bajos que los del esqueleto apendicular (Figura 4), principalmente en lo que refiere a las vértebras y costillas, es necesario considerar que para esta medida no fueron consideradas las piezas dentarias individuales, algo que sí se hizo para el MNE y podría explicar parte de esta diferencia.
Tabla 2. Composición anatómica y Mínimo Número de Elementos (MNE) por unidad de análisis.
Table 2. Anatomical composition and Minimum Number of Elements (MNE) per unit of analysis.
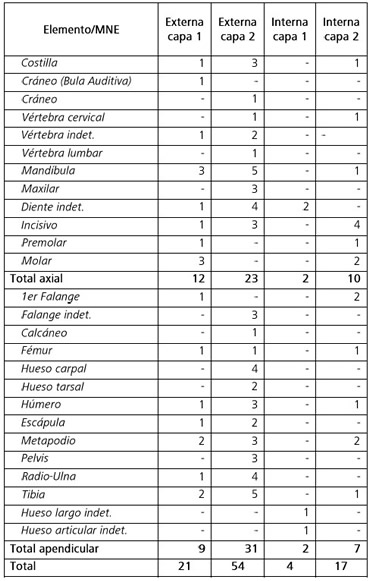
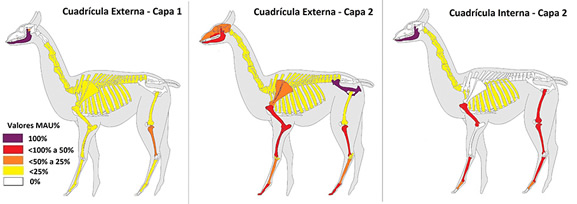
Figura 4. Representación del MAU% para Camelidae. Total de la muestra.
Figure 4. Representation of the MAU% for Camelidae. Sample total.
Para Camelidae, en total, se cuantificó un MNI de 18, de los cuales se pudo determinar el grupo etario de pertenencia de 17. Se observa un claro predominio de los individuos jóvenes, con un total de ocho individuos en toda la muestra. Le siguen los individuos neonatos (cuatro), los adultos (tres) y los viejos, con únicamente dos individuos (Tabla 3). En la capa 1 de la cuadrícula interna, dada la ausencia de partes diagnósticas, no se identificaron los grupos etarios.
Tabla 3. Grupos etarios de Camelidae por unidad de análisis.
Table 3. Age groups of Camelidae by unit of analysis.

En lo refiere al resto de los taxones y considerando su baja representación, la representación esqueletaria es bastante equilibrada, habiendo especímenes pertenecientes al esqueleto axial y apendicular en todos los casos. Los grupos etarios, por otro lado, solo pudieron ser identificados en un espécimen de Bos sp., el cual corresponde a un metapodio perteneciente a un individuo joven o novillo.
Los perfiles de meteorización presentes en la muestra son bastante heterogéneos, sobre todo en las unidades de análisis de la cuadrícula externa. En estos se observa la presencia de todos los estadios en proporciones similares. En el caso de la cuadrícula interna, la capa 1 presenta un predominio del estadio 5, observándose el patrón inverso en la capa 2 (Figura 5).
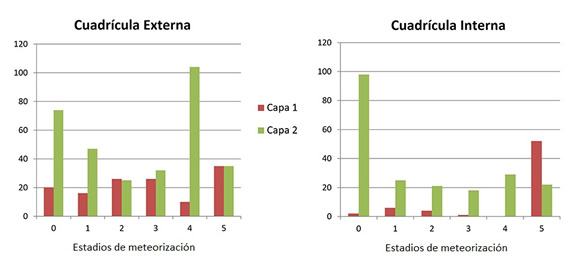
Figura 5. Meteorización por unidad de análisis. Total de la muestra.
Figure 5. Weathering by unit of analysis. Sample total.
Se observan patrones similares en el resto de las variables tafonómicas consideradas, habiendo mayores evidencias de acción de agentes bióticos y abióticos en los conjuntos del exterior (Tabla 4), aunque esta sea bastante reducida en el total de la muestra (únicamente un 7%). Resalta la acción de carnívoros, la cual predomina en la cuadrícula externa y es bastante intensa en varios de los especímenes. Esto último se identificó a partir de la presencia de varias marcas de distintos tipos en un mismo espécimen (tanto de Camelidae como de Ovicápridos) (Figura 6).
Tabla 4. Acción de agentes tafonómicos y antrópicos por unidad de análisis.
Table 4. Action of taphonomic and anthropic agents per unit of analysis.


Figura 6. Marcas de carnívoro. Camelidae: a) mascado y ahuecado sobre maxilar, b) hoyos, mascado y ranurado sobre calcáneo, c) hoyos sobre tibia. Ovicáprido: d) hoyos, mascado y ahuecado sobre escápula, e) hoyos, mascado y ahuecado sobre vértebra lumbar. Lagidium: f) hoyos sobre hueso largo.
Figure 6. Carnivore markings. Camelidae: a) chewed and hollowed on maxilla, b) pits, gnawed and scoring on calcaneus, c) pits on tibia. Ovicáprido: d) pits, gnawed and furrowing on the scapula, e) pits, gnawed and furrowing on lumbar vertebra. Lagidium: f) pits on long bone.
De los especímenes identificados a nivel taxonómico únicamente el 3,6% (29 especímenes) presenta marcas antrópicas. Aquellas identificadas en especímenes de Camelidae son principalmente de corte y se encuentran casi en su totalidad en el esqueleto apendicular, con excepción de una costilla y dos fragmentos de pelvis provenientes de los conjuntos de la capa 2 externa.
El predominio de las marcas de corte por sobre otros de tipos de marcas antrópicas es notorio a lo largo de toda la muestra, habiéndose identificado únicamente un espécimen con evidencia de machacado y otro con evidencias de percusión. Todas las marcas de corte identificadas se orientan de manera transversal al hueso, con excepción de un caso en el que se orientan de manera longitudinal. A su vez, todas las marcas se ubican sobre las epífisis de los huesos largos, con la excepción de cinco casos en los que se encuentran en la base o rodeando a la epífisis, y 18 en los que se distribuyen por el sector medio de la diáfisis (Figura 7). Del resto de los taxones, los únicos especímenes que presentan marcas antrópicas son uno de Ovicáprido y uno del género Lagidium. En ambos casos se trata de marcas de corte sobre una vértebra y un hueso largo respectivamente.

Figura 7. Marcas antrópicas de corte en Camelidae. a) Borde superior de escápula. b) Epífisis distal de húmero. c) Diáfisis de falange.
Figure 7. Anthropic cut marks in Camelidae. a) Upper edge of scapula. b) Distal epiphysis of humerus. c) Diaphysis of phalanx.
Se identificaron distintos tipos de fractura en 144 de los especímenes identificados (18%), siendo los ovicápridos el único taxón que no presenta esta modificación. Predomina la factura longitudinal, seguida por la espiral y la transversal (Tabla 2). Con respecto a la asociación de la fractura con marcas antrópicas, esta se identificó en dos casos, ambos correspondientes a especímenes de Camelidae. En el primero se observaron marcas de percusión y fractura longitudinal y espiral sobre una tibia, mientras que el segundo corresponde a un metapodio con marcas de machacado asociadas con fractura longitudinal. Por otro lado, también se ha identificado asociación de fractura con marcas de carnívoro en 15 especímenes.
Por último, se identificaron evidencias de termoalteración en un total de 195 especímenes (10% de la muestra total). El 47% se halló en los conjuntos de la cuadrícula externa, habiendo cantidades similares en ambas capas. La cuadrícula interna concentra el 53% y la mayor cantidad se acumula en la capa 2. Del total de especímenes, 59 se encuentran quemados y 136 calcinados, encontrándose estos últimos principalmente en la capa 2 interna y la capa 1 externa. Del total de especímenes termoalterados, 91 corresponden a especímenes identificados a nivel taxonómico y anatómico y, el resto (82), a especímenes no identificados y astillas. La mayoría de los especímenes identificados afectados corresponden a la clasificación Artiodactyla (65%), seguidos por el taxón Camelidae (31%).
Ningún espécimen de Bos sp. presenta evidencias de termoalteración. Por otro lado, los dos especímenes de Ovicáprido identificados en la capa 2 interna se encuentran afectados, siendo los únicos especímenes de este taxón que presentan termoalteración. Uno de ellos se encuentra quemado y el otro calcinado. Por último, Lagidium sp. presenta un espécimen quemado proveniente de la misma unidad de análisis.
Discusión
Procesos tafonómicos
Las diferencias en los perfiles de meteorización entre el interior y el exterior, así como la incidencia diferencial de la acción de distintos agentes tafonómicos, da cuenta de una preservación diferencial de los conjuntos óseos. En la cuadrícula interna, el predominio del estadio 1 y una baja representación de los estadios más avanzados (3 a 5) en la capa 2 podría estar indicando un entierro rápido de los especímenes óseos (Behrensmeyer, 1978), mientras que en la capa 1, el predominio del estadio 5 podría explicar la notable diferencia en la cantidad de especímenes entre esta unidad de análisis y la subyacente (la capa 1 contiene 104 especímenes y la capa 2, 522).
En la cuadrícula externa, por otro lado, la presencia de perfiles de meteorización heterogéneos en ambas capas podría estar indicando una acumulación atricional de restos óseos (Behrensmeyer, 1978), lo que explica el hecho de que especímenes pertenecientes a un mismo taxón (Camelidae) se encuentren en estadios de meteorización sumamente variados.
Con respecto a la acción de agentes tafonómicos bióticos y abióticos, si bien en el total de la muestra su incidencia es muy baja, se observa que los conjuntos de la cuadrícula externa se encuentran mucho más afectados que los del interior del recinto, sobre todo en lo que respecta a la acción de carnívoros, la cual puede deberse a una mayor exposición de los huesos depositados en el exterior.
Camélido
Como fue mencionado anteriormente, los camélidos dominan la muestra de manera notoria. La presencia en proporciones similares del esqueleto axial y apendicular en todas las unidades de análisis, así como la presencia de huesos de las articulaciones de las extremidades, estaría indicando que los esqueletos ingresaron completos al sitio. Esto da cuenta de que el lugar de matanza y procesamiento habría sido el mismo sitio o habría estado lo suficientemente cerca como para que los esqueletos pudieran ser descartados en el mismo lugar. Este patrón suele observarse, dentro del sistema de asentamiento pastoril, en las bases residenciales, sitios donde suelen llevarse a cabo estas actividades (Yacobaccio, 2007)
Si bien el MAU% muestra un predominio de los huesos apendiculares, es muy probable que esto esté ligado a la acción de procesos tafonómicos sobre huesos del esqueleto axial, tales como vértebras o costillas. La presencia de marcas de carnívoros sobre varios especímenes permite pensar en la incidencia de perros domésticos sobre el conjunto siendo un aspecto que podría estar sesgando la representación de huesos pequeños (Payne y Munson, 1985; Yacobaccio, 2007), aunque la misma también podría verse afectada por el accionar de zorros transportando estos restos óseos a sus madrigueras (Mondini, 1995). Sobre todo, considerando que los elementos de mayor tamaño, como la mandíbula y la pelvis, se encuentran representadas en porcentajes bastante altos dentro del conjunto. Dado el tamaño reducido del área donde se encuentran las cuadrículas (menor a los 4 m2) y su ubicación en un sector de difícil acceso, es poco probable que la muestra se encuentre sujeta a procesos destructivos tales como el pisoteo, sobre todo de animales.
Considerando que solo en capa 2 de la cuadrícula externa se recuperaron los restos de 11 individuos resulta notable la abundancia de ejemplares y variedad de taxones en relación al tamaño muestral y superficie excavada. La concentración de ejemplares en esta unidad de análisis sustenta, además, la posibilidad ya mencionada de que este conjunto corresponda a una acumulación generada por un descarte progresivo de restos de camélido. En las otras unidades de análisis, la cantidad de individuos representada es mucho menor y se observan principalmente restos de camélidos jóvenes y neonatos, grupos etarios con huesos de tamaño reducido cuya representación podría estar sesgada por las razones mencionadas más arriba.
En lo que respecta a los grupos etarios, distintos estudios etnoarqueológicos realizados en la región de la Puna plantean que la preponderancia de individuos adultos y viejos en los patrones de matanza está generalmente vinculado con el desarrollo de una estrategia destinada al transporte y la producción de fibra, mientras que una destinada al consumo de carne determinará que se priorice la matanza de animales jóvenes. Sin embargo, rara vez se observa el uso de una única estrategia y hay distintos factores que pueden afectar la composición de los grupos etarios en el registro arqueológico (Madero, 2000; Yacobaccio, 2007). Entre ellos se encuentran el tamaño del rebaño (la cantidad de animales disponibles y la posibilidad de consumirlos sin ejercer presión sobre el rebaño) y la disponibilidad de ganado menor como estrategia de evitación del riesgo (Madero, 2000).
Dado el predominio de individuos jóvenes y neonatos, es posible que los conjuntos de Antigal Laguna estén reflejando una estrategia de aprovechamiento de camélidos destinada al consumo de carne, algo observado en otros sitios arqueológicos de la zona (Mercolli, et al., 2018). A su vez, la presencia de restos de ovicápridos en el registro con evidencias de consumo permite pensar en la posibilidad de que estos animales estén jugando un rol complementario. Esto se observa hoy en día como estrategia de reducción de la presión sobre los camélidos frente a situaciones de riesgo ambiental y disminución del tamaño del rebaño (Madero, 2000).
Si bien la presencia de marcas antrópicas se observó únicamente en el 7% del total de especímenes de Camelidae, es relevante remarcar que esta son epifenómenos del procesamiento de los animales, por lo que no necesariamente se producen durante este proceso (Lyman, 1994). En la muestra, la presencia de marcas transversales ubicadas en las falanges y diáfisis de los metapodios identificadas en los conjuntos de la cuadrícula externa permite pensar que son producto de actividades de cuereo (Binford, 1981; Reitz y Wing, 1999). El resto de las marcas identificadas en la muestra son mayoritariamente de corte y parecen estar asociadas a actividades de carneo, dado que en varios casos se encuentran agrupadas, son en general cortas, se ubican principalmente en la metáfisis y se orientan de manera transversal al eje longitudinal de los huesos (Binford, 1981). La mayoría se encuentran en los huesos largos, salvo unas pocas excepciones ubicadas en la pelvis, costillas y escápula.
Con respecto a la fractura, se plantea que los patrones observados son producto de actividades intencionales, dado que es poco probable que los conjuntos se hubieran visto afectados por procesos tales como el pisoteo. Principalmente porque el área excavada se encuentra en un sector reducido y poco transitable. La cuadrícula externa se ubica en un pequeño espacio entre los recintos, menor a los 3m de ancho y adyacente a una terraza elevada de ignimbrita, lo que dificulta el tránsito de animales. Lo mismo ocurre en el caso de la cuadrícula interna, dado el pequeño tamaño del recinto (2m x 2m), área donde a lo sumo es posible considerar el pisoteo por parte de seres humanos.
Considerando ahora la fractura, es posible que en la muestra estén representadas tanto la producida por carnívoros como por seres humanos. Con respecto a la primera, los especímenes en el total de la muestra que presentan tanto marcas de dientes como fractura son 15, de los cuales ocho presentan las marcas de carnívoro sobre la diáfisis, en algunos casos incluso bordeando el perfil de la fractura, por lo que esta podría asociarse con la acción de estos animales (Mengoni Goñalons, 2010). Esto podría repetirse en los conjuntos provenientes de la cuadrícula externa, donde la acción de estos animales se encuentra mucho más representada.
Por último, del total de la muestra, el 10% presenta evidencias de termoalteración y, en el caso puntual del taxón Camelidae, el 8%. De todas estas estructuras de combustión presentes en la excavación, las que parecerían estar asociadas con restos faunísticos termoalterados corresponden a las que se encuentran en la capa 2 de la cuadrícula interna, dado que esta unidad de análisis concentra casi la totalidad de los especímenes de la cuadrícula que presentan esta alteración. A su vez, el amplio predominio de especímenes calcinados permite pensar que los restos óseos estuvieron expuestos a altas temperaturas durante bastante tiempo, por lo que es posible que los huesos hayan sido descartados dentro de los fogones o utilizados como combustible (Reitz y Wing, 1999).
Por otro lado, la mayor cantidad de restos termoalterados de la cuadrícula externa se encuentra en la capa 1, predominando los restos calcinados por sobre los quemados. En la capa 2 ocurre lo inverso. La cantidad de especímenes termoalterados es mucho menor y los que se encuentran quemados superan en número a los calcinados, por lo que es posible que los restos óseos sean producto de una limpieza de fogón, sobre todo en el caso de la capa 1.
A grandes rasgos es posible plantear que los camélidos identificados en el registro faunístico de Antigal Laguna son producto del procesamiento y descarte en el sitio. Las proporciones en las que se encuentran representados estos animales reflejan los patrones esperables en sitios arqueológicos pertenecientes a grupos de pastores en esta región, en los cuales suele predominar el taxón Camelidae, en detrimento de los ovicápridos (Madero, 2000).
Dentro de los sitios que estructuran el sistema de asentamiento pastoril andino, es probable que el sitio Antigal Laguna corresponda a una base residencial o “Casa de Campo”, dada la gran cantidad y variedad de estructuras que lo componen (Göbel 2002; Yacobaccio 2014). Esto también explicaría la enorme cantidad de restos faunísticos descartados en el sitio y la presencia del procesamiento de los esqueletos completos, sin mediar un traslado preferencial de partes esqueletarias (Yacobaccio 2007).
Otros taxones
En el caso de los ovicápridos, si bien los especímenes son muy pocos, la presencia de marcas antrópicas en una vértebra y termoalteración en dos fragmentos del arco cigomático permite pensar que los restos de estos animales son producto del consumo humano. A su vez, dado que es probable que las unidades de análisis correspondan a varios momentos distintos de descarte y que estos animales están representados en tres de las cuatro analizadas, se plantea que los restos posiblemente hayan sido consumidos en el sitio y de manera ocasional.
Sin embargo, no se puede descartar que el conjunto haya sido afectado por procesos tafonómicos, principalmente por carnívoros. La presencia de hoyos, mascado y ahuecado en dos de los cuatro especímenes de ovicápridos da cuenta de una acción sumamente intensa por parte de estos animales y, como fue mencionado más arriba, los huesos pequeños son más susceptibles de romperse (Payne y Munson, 1985; Yacobaccio, 2007) o ser acarreados por carnívoros más pequeños (Mondini, 1995), por lo que es posible que su presencia se encuentre sesgada.
De los pocos sitios que presentan restos de fauna europea en el registro en la región de la Puna, existen algunos que cuentan con material faunístico de cabras y ovejas en contextos domésticos, tanto en la Puna jujeña (Nielsen et al., 2015; Yacobaccio, comunicación personal) como en la catamarqueña (Quiroga 2015). La presencia de estos animales en el registro arqueológico de la Puna permite dar cuenta de que, si bien los camélidos siguen predominando en la dieta de los pastores andinos luego de la conquista, el ingreso de los ovicápridos a los rebaños habría implicado su inclusión en el consumo cotidiano, aunque no con la misma intensidad que las llamas. Esto podría deberse a su valor comercial en los mercados coloniales.
Los bóvidos, por otro lado, representan el 4% de los especímenes identificados y su distribución en las unidades de análisis es igual a la de los ovicápridos, encontrándose en todas las capas menos en la capa 1 interna. Se observa la presencia de fractura espiral y longitudinal en dos especímenes, probablemente intencional y antrópica, salvo por uno de los huesos (fragmento de radio-ulna proveniente de la capa 2 externa) en el que la fractura se encuentra acompañada por marcas producidas por carnívoro sobre la diáfisis. A diferencia de lo que ocurre con los camélidos y ovicápridos, los huesos de Bos sp. son mucho más grandes y densos, por lo que es difícil pensar en la posibilidad de acarreo por carnívoros menores y es más probable que hayan sido afectados por la acción de perros.
Las evidencias indicarían que los restos identificados en Antigal Laguna fueron consumidos y descartados en el sitio, sobre todo considerando la presencia de un individuo joven o novillo probablemente destinado a la obtención de carne, actividad que suele implicar el sacrificio de individuos en este rango etario (McGrory et al., 2012). Ahora bien, como fue mencionado antes, los especímenes de este taxón son muy pocos, por lo que sería necesario contar con una muestra mayor para poder derivar estas hipótesis con mayor seguridad.
En las fuentes históricas estos animales aparecen fuertemente vinculados al mercado colonial y siendo trasladados a través de la Puna por caminos de arriería para abastecer los centros mineros (Conti y Sica, 2011; Noli, 1998). Estas actividades tienen su correlato arqueológico en sitios como Casas Quemadas (Vaquer 2016) o Pan de Azúcar (Angiorama et al. 2015). Las menciones a la cría y consumo dentro de los circuitos pequeños y domésticos de estos animales en los trabajos realizados a partir de fuentes documentales son casi inexistentes.
Sin embargo, existen algunos contextos domésticos que dan cuenta de la presencia de bóvidos en actividades vinculadas con la vida cotidiana de los pastores, más que relacionados con actividades comerciales o productivas, como Punta de la Peña 3 (Quiroga 2015) en la Puna catamarqueña y San Lorenzo 1 (Nielsen et al. 2015), en la Puna jujeña. Antigal Laguna podría estar dando cuenta de una situación similar.
Por último, se destaca la presencia de dos ejemplares de Lagidium sp. con evidencias de procesamiento antrópico (marcas de corte, fractura y termoalteración). El consumo de estos animales por parte de los grupos humanos en la Puna tiene una larga historia, habiéndose registrado desde el Holoceno temprano (Yacobaccio y Morales, 2011) hasta el tardío, permaneciendo en la dieta incluso luego de la domesticación de los camélidos y la instauración de una economía pastoril (Olivera, 1997; Yacobaccio et al., 1998). Es por ello que no sorprende la presencia de este animal en el registro, dado que puede ser producto de un encuentro fortuito y de caza para su consumo. Nuevamente, la presencia de marcas de carnívoro sobre uno de los ejemplares permite pensar que la acción de estos agentes podría incidir en su baja representación.
Conclusiones
El objetivo principal que guio este trabajo fue el de indagar en las características que tuvo el proceso de introducción de las especies domésticas europeas en los rebaños de las poblaciones que habitaron el valle del río Barrancas entre los siglos XVII a XVIII. Se buscó conocer las estrategias de manejo para las distintas especies, las diferencias con los rebaños tradicionales de camélidos, cuáles fueron los usos y para qué fueron destinadas, de manera de poder evaluar no solamente cómo estos animales se incorporaron la vida cotidiana de los pastores, sino también de qué manera afectaron su participación en el contexto político y económico instaurado tras la conquista europea.
A nivel regional, de los pocos sitios arqueológicos en la Puna que cuentan con fauna europea en su registro, algunos presentan contextos similares al observado en Antigal Laguna. Tal es el caso de Punta de la Peña 3 (Quiroga, 2015) en la Puna catamarqueña y San Lorenzo 1 (Nielsen et al., 2015) y Puesto Demetria (Yacobaccio, comunicación personal) en la Puna jujeña. Esto sitios presentan restos de vacas y ovicápridos en contextos de tipo doméstico mostrando una continuidad en la ocupación y en los modos de vida con respecto a momentos prehispánicos.
Si bien Antigal Laguna presenta características que permiten plantear que se trata de un sitio doméstico, las evidencias a nivel regional permiten derivar hipótesis sobre los usos que se les dieron a las especies europeas presentes en el sitio. De acuerdo con lo planteado en las fuentes históricas, las especies europeas destinadas a la actividad ganadera eran sumamente valiosas dentro del sistema económico colonial, dado que eran destinadas principalmente al abastecimiento de los centros mineros. Los animales eran trasladados desde sus centros de crianza, a través de los distintos caminos de arrieros que atravesaban la Puna, hacia Potosí o centros mineros de menor escala. Sitios como Pan de Azúcar, centro minero de pequeña escala (Angiorama et al., 2015), y Casas Quemadas, paradero de arrieros (Vaquer, 2016), reflejan estas actividades en el registro arqueológico. La baja representación de Bos sp. y ovicápridos en el registro faunístico de Antigal Laguna puede estar vinculada, por un lado y como fue mencionado más arriba, a un consumo ocasional por parte de los habitantes de la localidad. Sin embargo, también permite preguntarse acerca de la posibilidad de que estuvieran destinadas a otro tipo de actividades, tales como su inserción en el circuito comercial de la Puna, dando cuenta de su baja representación en sitios de tipo domestico como Antigal Laguna.
En la actualidad, los rebaños de la Puna son de carácter mixto y las especies europeas (sobre todo cabras y ovejas) están completamente incorporadas, forman una parte estructural del sistema de pastoreo altoandino y proveen diversos recursos tales como carne, leche y lana. A su vez, el propio sistema de pastoreo debió conjugar las necesidades de los camélidos con las de los animales introducidos de manera que su estructura pueda responder a todas ellas (Yacobaccio et al., 1998).
Para los bóvidos, esta adaptación al ambiente árido del altiplano fue más compleja. La posibilidad de mantener al ganado vacuno depende de la existencia de campos de gran superficie, con abundante agua y pasturas, y su tenencia está más vinculada a cuestiones de estatus que a un uso concreto de los animales (Califano y Echazu, 2010).
Es probable que los patrones de manejo de las distintas especies en la actualidad no difieran demasiado de los llevados a cabo en el pasado. Esto es, por un lado, por la rapidez con la que se dispersaron e incorporaron las especies europeas a la cotidianeidad de las poblaciones locales. Por otro, porque el sistema de pastoreo, tanto en su organización general, como en las relaciones sociales que se estructuran en su entorno no se habrían visto tan afectadas por la conquista europea, como sí lo fueron sectores más fértiles y accesibles del NOA, como la Quebrada de Humahuaca y las zonas más bajas (Aramendi, 2016; Palomeque, 2000).
El registro faunístico del sitio Antigal Laguna parecería ajustarse a los patrones de consumo esperables para las distintas especies que componen la muestra, de acuerdo con lo planteado desde las fuentes históricas y desde la evidencia arqueológica previa, sobre todo en el caso de los camélidos y los ovicápridos. Sin embargo, la información de la que disponemos acerca del ingreso de estas especies en las economías pastoriles de la Puna, el manejo que se le dio a cada una y los roles que cumplieron, aún es muy escasa. Las fuentes disponibles para regiones aisladas de los centros de poder y de difícil accesibilidad son pocas, al igual que las investigaciones arqueológicas que aborden esta problemática, sumado a la poca presencia de estos animales en el registro. Sin embargo, los datos disponibles hasta ahora constituyen un punto de partida sumamente importante para responder estas preguntas.
El análisis del material faunístico obtenido de las excavaciones del sitio arqueológico Antigal Laguna permite evaluar, de manera preliminar, cómo se dieron estos procesos en la localidad de Barrancas, área donde el pastoreo parece constituir la actividad económica principal por lo menos desde hace 3200 años (Yacobaccio et al., 2018) y que continúa en la actualidad. Esto constituye un pequeño aporte a una problemática que es de suma importancia para comprender, por un lado, los profundos cambios que sufrió la organización social de las poblaciones que habitaban la región durante los momentos posteriores a la conquista europea. Por otro, para comprender de qué manera se constituyó la estructura del pastoreo andino actual, estrategia económica principal en la mayor parte de la Puna argentina y cuya configuración responde no solamente al aprovechamiento de los recursos que los animales puedan brindar, sino también a las variaciones ambientales propias de esta región y que requieren el desarrollo de estrategias que permitan hacer frente a situaciones de riesgo.
Ciudad de Buenos Aires, 17 de Agosto de 2021
Agradecimientos
Este trabajo se deprende de mi tesis de licenciatura. Los datos analizados en este trabajo se obtuvieron en el marco de un proyecto UBACyT, desarrollado entre 2014 y 2017. Agradezco a mi director, Hugo Yacobaccio por el apoyo y las correcciones, tanto de la tesis como de este trabajo. Agradezco también a los miembros del equipo y participantes del Proyecto Arqueológico Barrancas, en el marco del cual se realizó esta investigación. Por último, agradezco a los habitantes de Barrancas por la hospitalidad y por brindarnos la posibilidad de llevar a cabo este proyecto.
1. Adams, B., y Crabtree, P. (2012). Comparative Osteology. A Laboratory and Field Guide of Common North American Animals. Academic Press.
2. Angiorama, C., Becerra, M. F., y Pérez Pieroni, M. J. (2015). El mineral de Pan de Azúcar. Arqueología histórica de un centro minero colonial en la Puna de Jujuy (Argentina). Chungara, Revista de Antropología Chilena, 47(4), 603–619. https://doi.org/10.4067/S0717-73562015005000045
3. Aramendi, B. (2016). El distrito de la Puna y su primer subdelegado. Intendencia de Salta del Tucumán, 1784-1795. Fronteras de La Historia, 22(1), 12–37. https://doi.org/10.22380/20274688.12
4. Assadourian, C. S. (1982). El sistema de la economía colonial. Mercado interno, regiones y espacio económico. IEP ediciones.
5. Barrientos, G., Goñi, R., Zangrando, A., Del Papa, M., García Guraieb, S., Arregui, M. J., y Negro, C. (2007). Human taphonomy in Southern Patagonia: a view from the Salitroso Lake Basin (Santa Cruz, Argentina). Taphonomy and Zooarchaeology in Argentina, 1601(1426), 187–201.
6. Behrensmeyer, A. K. (1978). Taphonomic and ecologic information from bone weathering. Paleobiology, 4(2), 150–162.
7. Binford, L. R. (1981). Bones. Ancient men and modern myths. Academic Press.
8. Browman, D. L. (1987). Agro-pastoral risk managment in the central Andes. En B. L. Isaac (Ed.), Research in Economic Anthropology (Vol. 8, pp. 171-200). JAI Press Inc.
9. Cabrera, A. L. (1976). Regiones Fitogeográficas Argentinas. ACME.
10. Califano, M. y Echazu, F. (2010). La diversidad de los sistemas ganaderos campesinos en la Puna jujeña. Determinación en el marco de un proyecto de extensión. Aportes teórico-metodológicos para el trabajo de Extensión Rural. La dinámica socio-económica, ambiental y organizacional. Trabajo presentado en XV Jornadas Nacionales de Extensión Rural, Potrero de los Funes, San Luis, Argentina.
11. Capparelli, A., Lema, V., Giovannetti, M., y Raffino, R. (2005). The introduction of Old World crops (wheat, barley and peach) in Andean Argentina during the 16th century AD: archaeobotanical and ethnohistorical evidence. Vegetation History and Archaeobotany, 14(4), 472–484. https://doi.org/10.1007/s00334-005-0093-8
12. Conti, V., y Sica, G. (2011). Arrieros andinos de la colonia a la independencia. El negocio de la arriería en Jujuy, Noroeste Argentino. Nuevos Mundos, Mundos Nuevos, 20–37. https://doi.org/10.4000/nuevomundo.60560
13. Cornaglia Fernandez, J. (2012). El carbonato de calcio y sus implicancias en el análisis de los conjuntos arquefaunísticos. El caso de Laguna El Doce (Departamento General Lopez, Provincia de Santa Fe). Revista Del Museo de Antropologia, 5(1), 185–194. https://doi.org/10.31048/1852.4826.v5.n1.9138
14. Göebel, B. (2002). La arquitectura del pastoreo: uso del espacio y sistemas de asentamiento en la Puna de Atacama (Susques). Estudios Atacameños, 23, 53-73. DOI:10.4067/S0718-10432002002300005
15. Gil Montero, R. (2009). Mountain pastoralism in the Andes during colonial times. Nomadic Peoples, 13(2), 36–50. https://doi.org/10.3167/np.2009-1302203
16. Hongo, H. (1998). Patterns of animal husbandry in central Anatolia in the second and first millenia BC: Faunal remains from Kaman-kalehöyük, Turkey. En H. Buitenhuis, L. Bartosiewicz, & A. M. Choyke (Eds.), Achaeozoology of the Near East III. Procedings of the third international symposium on the archaeological of southwestern Asia and adjacent areas (pp. 255–275). ARC (Publicatie 18).
17. Hopkins, C. E., Hamm, T. E., y Leppart, G. L. (1970). Atlas of goat anatomy. Part I: Osteology. Department of the Army.
18. Lyman, R. L. (1994). Vertebrate Taphonomy. Cambridge University Press.
19. Madero, C. M. (2000). Modelo etnoarqueológico de estructura etaria en contextos pastoriles actuales de la Puna de Jujuy. Cuadernos Del Instituto Nacional de Antropología y Pensamiento Latinoamericano, 19, 285–399.
20. McGrory, S., Svensson, E., Götherström, A., Mulville, J., Powell, A. J., Collins, M. J. y O’Connor, T. P. (2012). A novel method for integrated age and sex determination from archaeological cattle mandibles. Journal of Archaeological Science, 9(10), 3324-3330. DOI: 10.1016/j.jas.2012.05.021
21. Mengoni Goñalons, G. L. (1980). Notas zooarqueológicas I: Fractura en huesos. En Actas del VII Congreso Nacional de Arqueología (pp. 87–91). Centro de Estudios de Arqueología Montevideo.
22. Mengoni Goñalons, G. L. (1999). Cazadores de guanacos de la estepa patagónica. Colección Tesis Doctorales. Sociedad Argentina de Antropología.
23. Mengoni Goñalons, G. L. (2010). Zooarqueología en la práctica: algunos temas metodológicos. Xama, 19(23), 83-113.
24. Mercolli, P., Mamani, H. y Basso, D. M. (2018). Resultados del análisis de la muestra arqueofaunística del sitio Coch-1.1 (Cochinoca, Puna de Jujuy, Argentina). Cuadernos del Instituto Nacional de Antropología y Pensamiento Latinoamericano, 6(2), 39-54.
25. Mondini, M. (1995). Artiodactyl prey transport by foxes in Puna rock shelters. Current Anthropology, 36(3), 520-524.
26. Morales, M. R., Bustos, S., Oxman, B. I., Pirola, M., Tchilinguirián, P., Orgeira, M. J., y Yacobaccio, H. D. (2018). Exploring habitat diversity of mid-holocene hunter-gatherers in the South-Central Andes: Multi-proxy analysis of Cruces Core 1 (TC1), Dry Puna of Jujuy, Argentina. Journal of Archaeological Science: Reports, 18(January), 708–721. https://doi.org/10.1016/j.jasrep.2017.07.010
27. Nielsen, A. E., Angiorama, C., Maryñanski, J., Avila, F., y López, M. L. (2015). Paisajes prehispánicos tardíos en San Juan Mayo (Frontera Argentina-Bolivia). Arqueología, 21, 33–65.
28. Noli, E. S. (1998). Algarrobo, maíz y vacas. Los pueblos indios de San Miguel de Tucumán y la introducción de ganados europeos (1600-1630 ). Mundo de Antes - Instituto de Arqueología y Museo (UNT), 1, 31–68.
29. Olivera, D. E. (1997). La importancia del recurso Camelidae en la Puna de Atacama entre los 10.000 y 500 años A.P. Estudios Atacameños, 14(14), 29-41. DOI: 10.2307/25674670
30. Otero, C., y Rivolta, M. C. (2015). Nuevas interpretaciones para la secuencia de ocupación de Tilcara (Quebrada de Humahuaca, Jujuy). Intersecciones En Antropologia, 16(1), 145–159.
31. Pacheco Torres, V. R., Altamiro, A. J., y Guerra Porras, E. S. (1986). The osteology of South American camelids. Institute of Archaeology, University of California.
32. Palomeque, S. (2000). El mundo indígena (siglos XVI -XVII). En Nueva Historia Argentina, (Tomo 2, pp. 87–145). Sudamericana.
33. Payne, P y Munson, P. (1985). Ruby and how many squirrels? The destruction of bones by dogs. En N. Fieller, D. Gilberston & N. Ralph (Eds.), Paleobiological Investigations. Research design, methods and data analysis, (pp. 31-39). Oxford British Archeologycal Reports.
34. Quiroga, L. (2015). Del páramo a la Puna. Textos y contextos arqueológicos para una descripción del paisaje altoandino en la gobernación del Tucumán. Corpus. Archivos Virtuales de La Alteridad Americana, 5(2), 1-21. https://doi.org/10.4000/corpusarchivos.1516.
35. Rabey, M. A., Rotondaro, R., y Fernandez Distel, A. (2008). Temas de Barrancas: Un pueblo de la Puna. Editorial Hanne.
36. Reitz, E. y Wing, E. S. (1999). Zooarchaeology. Cambridge University Press.
37. Samec, C. (2014). Ecología isotópica en la Puna seca argentina: un marco de referencia para el estudio de las estrategias de pastoreo en el pasado. Cuadernos del Instituto Nacional de Antropología y Pensamiento Latinoamericano. Series Especiales, 2(1), 61-85.
38. Sica, G. (2005). Maíz y trigo; molinos y conanas; mulas y llamas. Tierras, cambio agrario, participación mercantil indígena en los inicios del sistema colonial. Jujuy. Siglo XVII. En D. J. Santamaría (Ed.), Jujuy : arqueología, historia, economía, sociedad (pp. 106–124). Centro de Estudios Indígenas y Coloniales.
39. Vaquer, J. M. (2016). La ocupación colonial temprana (S . XVI y XVII) en Casas Quemadas (Cusi Cusi , Rinconada , Jujuy): primeras aproximaciones a las relaciones entre lo global y lo local. Revista de Arqueología Histórica Argentina y Latinoamericana, 10(2), 1–26.
40. Von Rosen, E. (1957). Restos de una cultura indígena. En Un mundo que se va (pp. 87–125). Instituto Miguel Lillo - Universidad Nacional de Tucumán.
41. Wheeler, J. C. (1982). Aging llamas and alpacas by their teeth. Llama World, 1(2), 12–17.
42. Yacobaccio, H. D. (2007). Andean camelid herding in the South Andes: ethnoarchaeological models for archaeozoological research. Anthropozoologica, 42(2), 143–154.
43. Yacobaccio, H. D. (2014). Pastoreo, movilidad y sequías. Cuadernos del Instituto Nacional de Antropología y Pensamiento Latinoamericano – Series Especiales, 2(1), 113-121.
44. Yacobaccio, H. D., Madero, C. M., Malmierca, M. P., & Reigadas, M. del C. (1998). Caza , domesticacion y pastoreo de camélidos en la Puna argentina. Relaciones de La Sociedad Argentina de Antropología, 22, 389–418.
45. Yacobaccio, H. D., & Morales, M. R. (2011). Ambientes pleistocénicos y ocupación humana temprana en la Puna argentina. Boletín de Arqueología PUCP, (15), 337–356.
46. Yacobaccio, H. D., Hoguin, R., Morales, M. R., Solá, P., Oxman, B. I., Samec, C. T., Pirola, M., Bustos, S., Merler Carbajo, J., Rouan Sirolli, M. y Kohan, P. (2018). Proyecto Barrancas: Informe de Avance Científico (Inedito).